 In acest tip de reactie elementul esential il formeaza populatia de limfocite T (timodependente) prezente in timus, zonele paracorticale ale ganglionilor limfatici si in sange, unde formeaza aproximativ 2/3 din totalul limfocitelor circulante. Aceste limfocite pot fi deosebite morfologic numai la microscopul electronic de cele de tip B, ele avand o membrana celulara neteda, fara vilozitati. Felul in care ele sunt sensibilizate (facute sa reactioneze specific) cu antigenul a fost mult timp de domeniul ipotezelor. Dificultatea venea de la faptul ca suprafata membranei lor nu prezenta locusuri de unire pentru nici o structura antigenica cunoscuta. Se stie ca limfocitele B au locusuri de legatura cu portiunea Fc a diverselor tipuri de imunoglobuline (G, M, D, A, E), ca si pentru lanturile usoare (k, K) si pentru fractiunea C3. Un numar variabil de limfocite T pot fixa pe suprafata lor astfel de structuri; aceasta se intampla in cursul a diverse reactii imunologice la care participa si populatia de limfocite. Fixarea de structuri straine limfocitului T in acest caz este secundara. De curand s-a aratat ca limfocitul T are si el in structura membranei locusuri formate endogen de celula, caracterizate prin structuri polipeptidice de aproximativ 75 000 daltoni, care exista ca dimeri si care reactioneaza cu diverse imunoglobuline (si cu diversi antigeni) de un tip special (cu lanturi grele, mono- sau dimeri, cu o parte constanta — C — speciala deosebita de a celorlalte imunoglobuline si care a facut ca imunoglobulinei respective sa i se dea un nume aparte, de lant greu t-Tau. O alta contributie recenta la cunoasterea celulei T este ca in acelasi timp cu recunoasterea si legarea de antigen (pe locusuri t), pentru a obtine un raspuns imunologic maximal din partea limfocitului, acesta trebuie sa recunoasca in structura antigenului tinta si antigene de histocompatibilitate majora. Astfel de locusuri antigenice speciale, care sa duca la sensibilizarea limfocitului T si amorsarea unei reactii de tip IV, se gasesc probabil in structurile complexe care induc hiperergie de tip celular (anumiti virusi si bacterii sau fungi, anumite complexe formate din haptene — substante chimice sau droguri — si proteine ale organismului respectiv sau in organe transplantate sau tumori).
In acest tip de reactie elementul esential il formeaza populatia de limfocite T (timodependente) prezente in timus, zonele paracorticale ale ganglionilor limfatici si in sange, unde formeaza aproximativ 2/3 din totalul limfocitelor circulante. Aceste limfocite pot fi deosebite morfologic numai la microscopul electronic de cele de tip B, ele avand o membrana celulara neteda, fara vilozitati. Felul in care ele sunt sensibilizate (facute sa reactioneze specific) cu antigenul a fost mult timp de domeniul ipotezelor. Dificultatea venea de la faptul ca suprafata membranei lor nu prezenta locusuri de unire pentru nici o structura antigenica cunoscuta. Se stie ca limfocitele B au locusuri de legatura cu portiunea Fc a diverselor tipuri de imunoglobuline (G, M, D, A, E), ca si pentru lanturile usoare (k, K) si pentru fractiunea C3. Un numar variabil de limfocite T pot fixa pe suprafata lor astfel de structuri; aceasta se intampla in cursul a diverse reactii imunologice la care participa si populatia de limfocite. Fixarea de structuri straine limfocitului T in acest caz este secundara. De curand s-a aratat ca limfocitul T are si el in structura membranei locusuri formate endogen de celula, caracterizate prin structuri polipeptidice de aproximativ 75 000 daltoni, care exista ca dimeri si care reactioneaza cu diverse imunoglobuline (si cu diversi antigeni) de un tip special (cu lanturi grele, mono- sau dimeri, cu o parte constanta — C — speciala deosebita de a celorlalte imunoglobuline si care a facut ca imunoglobulinei respective sa i se dea un nume aparte, de lant greu t-Tau. O alta contributie recenta la cunoasterea celulei T este ca in acelasi timp cu recunoasterea si legarea de antigen (pe locusuri t), pentru a obtine un raspuns imunologic maximal din partea limfocitului, acesta trebuie sa recunoasca in structura antigenului tinta si antigene de histocompatibilitate majora. Astfel de locusuri antigenice speciale, care sa duca la sensibilizarea limfocitului T si amorsarea unei reactii de tip IV, se gasesc probabil in structurile complexe care induc hiperergie de tip celular (anumiti virusi si bacterii sau fungi, anumite complexe formate din haptene — substante chimice sau droguri — si proteine ale organismului respectiv sau in organe transplantate sau tumori).
Mecanismele efectoare, cauzatoare ale starii de hiperergie, sunt multiple si se intrica; uneori au o intensitate slaba si efectul final este de anihilare, de distrugere a antigenului, jucand astfel un rol in apararea organismului (in imunitate), atunci cand antigenul este un agent agresor; alteori ele sunt de intensitate mare si, daca dureaza mai mult timp, au un efect distructiv asupra diverselor structuri ale organismului.
-Concomitent cu factorul de inhibare, limfocitele elibereaza in mediul inconjurator substante care activeaza mult metabolismul macrofagelor, care devin foarte active si agresive (suparate), putind fagocita si distruge mai eficient si rapid germenii ingerati, daca antigenul este germen, sau capata o actiune agresiva, distructiva asupra structurilor vecine zonelor in care se afla; aceste substante se numesc limfokine.
-Factorul chemotactic (de atragere) monocitar.
-Factorul reactiv cutanat care produce, daca reactia are loc in tegumente, permeabilitate crescuta capilara si exsudat (aparitia de edem).
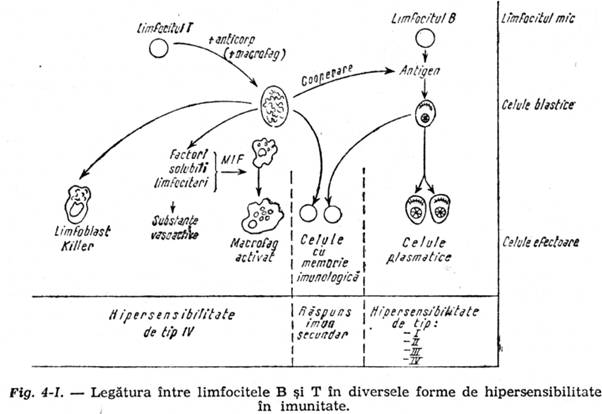
-Factorul mitotic care induce mitoze (transformarea blastica) in populatiile de celule limfatice nesensibilizate. in felul acesta creste numarul de celule active, atat limfocite T cat si B; celulele T active intervin favorizind si crescand actiunea de formare de anticorpi ai celulelor B (fenomen de cooperare) care, datorita factorului mitotic, sunt in faza activa, producand anticorpi specifici antiantigenul respectiv. Iata deci ca fenomenul de hiperergie celulara se complica cu cel de hiperergie umorala (fig. 4-1). Nu toate antigenele care induc alergie de tip IV induc si alergie umorala. Producerea excesiva de anticorpi serici specifici, la injectarea unui antigen, atunci cand antigenul este inglobat in reactiv Freund (suspensie de ulei in apa continand bacili Koch omorati), este datorita asocierii incitatiei antigenice pe limfocitul B in formarea de anticorpi, incitatie care dureaza mai mult timp, antigenul fiind inglobat intr-osolutie uleioasa greu rezorbabila, insa si actiunii de cooperare intre limfocitul T activat de antigenul din bacilul Koch, cu limfocitul B pe care il incita la producerea de anticorpi. Nu toate antigenele inglobate in reactiv Freund au acest dublu efect stimulator in producerea de anticorpi specifici. Cele care il au se numesc timodependente.
 Factorii amintiti induc, in zona in care se gasesc, o aglomerare mare de celule; dintre acestea, numai foarte putine sunt angrenate specific, cele mai multe fiind macrofage, mononucleare si limfoblasti aparuti sub influenta factorilor amintiti (efect de amplificare). Aspectul histologic al leziunii este caracteristic; initial, aglomerarea perivasculara de mononucleare, careia ii urmeaza exsudare intensa, cu o populatie formata in special din mononucleare si polinucleare. Acestea din urma migreaza rapid din zona leziunii, ramanand o populatie celulara formata aproape exclusiv din macrofage, monocite, limfoblasti si limfocite.
Factorii amintiti induc, in zona in care se gasesc, o aglomerare mare de celule; dintre acestea, numai foarte putine sunt angrenate specific, cele mai multe fiind macrofage, mononucleare si limfoblasti aparuti sub influenta factorilor amintiti (efect de amplificare). Aspectul histologic al leziunii este caracteristic; initial, aglomerarea perivasculara de mononucleare, careia ii urmeaza exsudare intensa, cu o populatie formata in special din mononucleare si polinucleare. Acestea din urma migreaza rapid din zona leziunii, ramanand o populatie celulara formata aproape exclusiv din macrofage, monocite, limfoblasti si limfocite.
In patologie, infectia tuberculoasa induce mecanisme patogenice de tipul hipersensibilitatii celulare. Aceasta este evidentiata de doua teste care au importanta si in diagnosticul bolii; intradermoreactia la tuber- culina, reactie imaginata de Mantoux, si transformarea blastica la P.P.D.
Reactia Mantoux, consta in injectarea intradermica a (1—2) unitati P.P.D. Daca subiectul este sensibilizat la Mycobacterium tuberculosis, dupa 24—48 ore la locul injectiei apare o zona de eritem si induratie (reactie intarziata), insotita sau nu de reactie regionala (iimiangita, adenopatie dureroasa regionala) si sistemica (febra).










































_13244247920.jpg)





















_13466647540.jpg)
_13655255750.jpg)
