Acest tip de reactie este caracterizat prin prezenta in sangele subiectului respectiv a unei mase de molecule proteice de felul IgG sau IgM. Acestea au o afinitate specifica (se comporta ca anticorpi) pentru anumite antigene, care fac parte fie din structura membranei celulelor libere circulante in sangele sau/si umorile organismului (hematii, trombocite, leucocite) sau a unor celule fixe apartinand diverselor organe sau tesuturi, fie ca fac parte din structura membranelor bazale vasculare. in unele cazuri anticorpii au afinitate specifica pentru structuri antigenice complexe, care numai in parte sunt proprii celulelor amintite sau membranelor bazale si care se fixeaza specific pe astfel de structuri. Este vorba de diverse substante introduse in organism de multe ori cu scop terapeutic. Astfel de substante se comporta in organism, la anume indivizi, ca haptene, in sensul ca se cupleaza cu o structura proteica, ce face parte din textura membranei celulare sau bazale, formand cu aceasta un complex cu proprietati antigenice care induc formarea de anticorpi specifici. In unele cazuri rolul de haptena il joaca diverse ma- cromolecule polizaharidice, rezultate cel mai adesea din distrugerea de germeni gramnegativi, de obicei in cadrul unor infectii fortuite.
Prezenta de anticorpi si antigene specifice creeaza in organismul respectiv o situatie critica; aceasta este caracterizata imunologic printr-o reactie antigen-anticorp in sensul ca anticorpul se fixeaza specific pe antigen, initiind o serie de tulburari in celula sau membrana bazala respectiva care tulbura, uneori grav, morfologia si functionalitatea acestora (de aici numele de reactie citotoxica), mergand pana la distrugerea structurii respective. Clinic aceste tulburari se caracterizeaza prin aparitia semnelor de boala care variaza, dupa structurile afectate (si dupa modul de aparitie a anticorpilor).
Citotoxicitatea sau afectarea membranei bazale indusa de complexele antigen-anticorp formate poate sa se produca prin diverse mecanisme:
a) Uneori complexele antigen-anticorp pot induce activarea secventiala (clasica) a componentelor complementului (C) seric, Ct> 4,2 apoi C3 care ramane fixat ferm pe celula (C3b) si induce activarea C5,6,7 care activeaza C8,9 complex care fixat pe membrana celulei are o actiune distructiva locala. Daca celula este fara nucleu (hematie, trombocit) posibilitatile ei de a repara o astfel de leziune sunt minime si celula sufera procesul de liza imuna, care duce in final la distrugerea ei. Este suficient un singur locul de activare a C pe suprafata hematiei pentru a o distruge. Daca celula care sufera agresiunea este nucleata, posibilitatile de reparatie cresc, asa ca sunt necesare, dupa tipul de celula, locusuri multiple de activare a C pentru a duce in final la liza imuna a celulei.
In cazurile in care complexul antigen-anticorp are in structura sa polizaharide, acesta poate activa C seric pe o alta cale (alterna); activand C3 care activeaza secvential C5,6, si in final C.s> 9 cu rolul aratat.
Iata deci cum un complex specific antigen-anticorp poate produce distrugeri grave celulare (sau de membrane bazale) printr-un mecanism nespecific de activare a complementului (C). Activarea C nu este totusi des intilnita in hiperergiile de tip II.
b) Fixarea C3b pe suprafata structurii agresionate induce si un alt mecanism — nespecific — de distrugere a structurii respective. Este fenomenul denumit imunoaderenta. Fagocitele au receptori pentru C3b, asa ca ele migreaza fixandu-se pe locusul cu C3b, fenomen care favorizeaza fagocitoza celulei respective.
c) Afluxul de fagocite si initierea fagocitozei sunt induse si de prezenta anticorpului pe structura celulei sau a membranei bazale agresionate. Anticorpul este o imunoglobulina G sau M care are in compozitia ei o parte denumita Fc; fagocitele au locusuri complementare acestei structuri cu care complexeaza, in felul acesta se initiaza afluxul de fagocite si procesul fagocitozei (fenomen denumit aderenta opsonica).
Factorul destructiv pentru celula agresionata nu consta numai in fagocitoza, ci si in faptul ca fagocitele acumulate local elibereaza enzime lizozomale puternic proteolitice; acest fenomen se produce cu intensitate mai mare in cazurile in care interactiunea antigen-anticorp are loc cu activarea clasica a complementului; activarea C3 duce la scindarea moleculei in doua parti: C3b de care s-a amintit si C3a denumita si anafila- toxina, cu actiune puternica vasodilatatoare si chemotactica (de atragere a leucocitelor). Activarea C5 duce la formarea a doua molecule: C5b si C-a. Aceasta din urma are si ea proprietati chemotactice si vaso- dilatatorii, inducand un influx crescut de leucocite in zona in care ea este prezenta.
d)Un alt mecanism de atac citotoxic este cel initiat de o populatie de celule care nu au proprietati fagocitante, denumite celulele K. Ele au un caracter morfologic limforeticular, uneori mieloid. Aceste celule nu sunt sensibilizate pentru complexele imune; ele au insa locusuri in membrana lor de reactie cu portiunea Fc a imunoglobulinei G, asa ca sunt atrase de astfel de structuri. Este cunoscut ca anticorpii specifici se fixeaza pe aceste structuri cu portiunea variabila, lasand libera portiunea denumita Fc. De aceasta se fixeaza celulele K si induc distrugerea celulei respective. Atunci cand celulele K se combina cu structuri antigen-anticorp din membrana bazala sau din endoteliul vascular, induc si un fenomen de aderenta plachetara cu agregarea trombocitelor si tromboza vasculara cu consecinta ei, ischemia si detractarea structurilor irigate de vasul respectiv. Populatiile de celule K sunt inca incomplet definite, ca origine, proprietati morfofunctionale sau ca modalitate de atac, care se banuieste ca este extracelulara si nefagocitica. Acest atac se intampla in special in cazuri de hipersensibilizare la paraziti mari sau tumori, sau in cazurile de transplant de organ.
e) In unele cazuri celulele K fixeaza pe suprafata lor complexe antigen-anticorp eliberate in sange din celulele agresionate si distruse. In aceste complexe imune uneori valentele anticorpului sunt insuficient saturate de antigen, ramanand in parte libere. Prin aceste valente libere de anticorp celula K este atrasa de o celula de distrus (celula tinta),
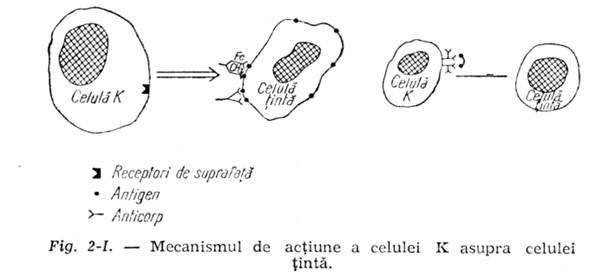 care are fixata pe suprafata ei complexe antigen-anticorp, avand insa uneori si structuri antigenice pe care nu s-au fixat anticorpi specifici (fig. 2-1). Unindu-se prin legatura valentei libere, anticorpul de pe ce¬lula K, cu antigenul liber de pe celula tinta, celula K o distruge, cum s-a aratat in paragraful anterior. Este vorba de o distrugere de asta data specifica a celulei tinta, de catre o celula K inarmata cu complexe imune antigen-anticorp. Acest fenomen se intampla in unele forme de hipersensibilizare la transplante de organe sau in tumori.
care are fixata pe suprafata ei complexe antigen-anticorp, avand insa uneori si structuri antigenice pe care nu s-au fixat anticorpi specifici (fig. 2-1). Unindu-se prin legatura valentei libere, anticorpul de pe ce¬lula K, cu antigenul liber de pe celula tinta, celula K o distruge, cum s-a aratat in paragraful anterior. Este vorba de o distrugere de asta data specifica a celulei tinta, de catre o celula K inarmata cu complexe imune antigen-anticorp. Acest fenomen se intampla in unele forme de hipersensibilizare la transplante de organe sau in tumori.
Efectele distructive induse de acest tip de hipersensibilizare vor fi in special severe atunci cand celula agresionata (celula tinta) este libera (hematii, trombocite sau leucocite) sau este vorba de un endoteliu vascular (de obicei renal sau pulmonar); efectele vor avea o pondere mult mai mica clinica atunci cand celula tinta face parte dintr-un parenchim compact.










































_13244247920.jpg)





















_13466647540.jpg)
_13655255750.jpg)
